À côté de la synapse, structure de communication la plus élaborée, il en existe d’autres et nous indiquerons à titre d’exemple les jonctions communicantes et les interactions par contact.
Synapse
La synapse est une structure spécialisée dans la transmission des informations. Elle peut se définir comme une zone spéciale de contact entre deux neurones ou entre un neurone et un élément non neuronal. Elle comprend un élément présynaptique, une fente et un élément postsynaptique.
La transmission synaptique comporte plusieurs étapes:
- Synthèse et stockage du médiateur dans l’élément présynaptique: le stockage s’effectue dans des vésicules intracytoplasmiques grâce à des transporteurs dont certains sont inhibés par un médicament qui n’est plus utilisé en thérapeutique, la réserpine.
- Libération du médiateur par l’élément présynaptique dans la fente synaptique: cette libération comporte la migration de la vésicule contenant le médiateur du cytoplasme jusqu’à la membrane plasmique, sa fusion avec cette membrane, enfin la libération du médiateur. Interviennent dans ces phénomènes le calcium, les canaux calciques voltage-dépendants, diverses protéines com-me les syntaxines, la synaptobrévine, la synaptotagmine. En général, il y a co-libération, c’est-à-dire libération concomitante de deux ou plusieurs messagers.
- Interactions du médiateur avec ses récepteurs spécifiques postsynaptiques (R1 et R2) et présynaptiques
- Recapture du médiateur par l’élément présynaptique. La recapture d’un médiateur est liée à un transport de sodium : soit le cotransport Na+/Cl (probablement deux Na+ pour un Cl–)soit à l’échange Na+/ K+ ou Na+ /OH–. Les transporteurs Na+/Cl– dépendants assurent d’une manière spécifique pour chacun d’eux la recapture de noradrénaline, de dopamine, de sérotonine, de GABA, de glycine ou de taurine et les transporteurs liés aux échanges Na+/ K+ ou Na+/ OH– assurent la recapture de glutamate et d’aspartate. Ces derniers transporteurs sont également présents au niveau des cellules gliales.
- Diffusion du médiateur, catabolisme, fixation non spécifique par les tissus.
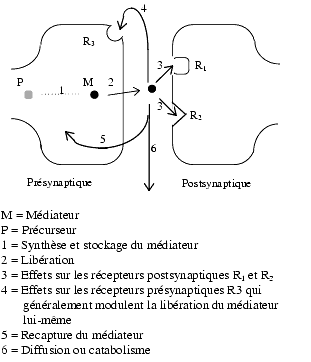 Schéma d’une synapse
Schéma d’une synapse- Effets résultant de l’interaction du médiateur avec un ou plusieurs types de récepteurs.
- postsynaptiques : leur stimulation entraîne des effets principaux qui dépendent du système de transduction (par exemple les protéines Gs et Gi) reliant le récepteur à l’effecteur final (par exemple une fibre lisse qui pourra se contracter ou se relâcher).
- présynaptiques : leur stimulation module la libération du médiateur lui-même. Ils sont appelés autorécepteurs car ils modulent la libération du médiateur qui les active. Certains inhibent la libération (par exemple le récepteur a2 pour la noradrénaline, D2/D3 pour la dopamine, M2 pour l’acétylcholine, 5-HT1D pour la sérotonine, H3 pour l’histamine) ; d’autres l’augmentent (par exemple le récepteur ß2 de la noradrénaline).
- Il existe aussi au niveau des terminaisons présynaptiques des hétérorécepteurs, ainsi appelés parce qu’ils modulent la libération du médiateur considéré tout en étant activés par d’autres médiateurs.
Synapses du système nerveux autonome
Le système nerveux autonome comprend le sympathique et le parasympathique, chacun formé d’une fibre présynaptique et d’une fibre postsynaptique séparées par un relais ganglionnaire et l’acétylcholine assure la transmission de l’information entre ces deux fibres. La fibre postsynaptique, du côté sympathique, libère de la noradrénaline, et du côté parasympathique, de l’acétylcholine. Le relais ganglionnaire parasympathique est situé à proximité immédiate des organes innervés par lui alors que le relais sympathique est situé à distance, plus près des centres. D’une manière générale l’innervation sympathique est plus largement distribuée dans l’organisme que l’innervation parasympathique. 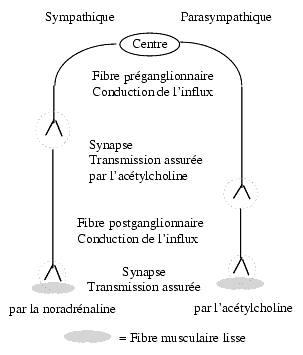
- La synapse assurant la transmission neuromusculaire (muscle strié) fonctionne à la manière d’un relais ganglionnaire, l’acétylcholine étant le médiateur.
Synapses du système nerveux central
A la simplicité du système nerveux autonome s’oppose la complexité du système nerveux central. Un cerveau comporte environ cent milliards de neurones et chaque neurone reçoit des dizaines de milliers de contacts synaptiques. Chaque neurone est sous l’influence simultanée de plusieurs médiateurs aux effets semblables ou opposés.
Les raisonnements que nous utilisons pour expliquer les effets d’un médicament sur le système nerveux central mettent l’accent sur telle ou telle propriété de ce médicament, mais il faut savoir que ce sont des simplifications extrêmes d’une réalité infiniment complexe.
Autres structures
Jonctions communicantes
La communication entre cellules contiguës peut aussi se faire par jonctions communicantes ou couplage jonctionnel. Les canaux jonctionnels sont constitués de connexines qui assemblées par six forment une structure tubulaire creuse appelée connexon (désigné par l’abréviation Cx suivie d’un chiffre exprimant le poids moléculaire en kilodalton). Lorsque les connexons de deux cellules contiguës s’alignent bout à bout ils forment un canal hydrophile qui, à l’état ouvert, permet le passage de cations, de métabolites, de messagers secondaires d’une cellule à l’autre et peut-être aussi de médicaments. Les zones de contact entre les membranes plasmiques sont extrêmement étroites et appelées «gap junction». Ces jonctions sont des structures dynamiques passant de l’état ouvert à l’état fermé; leur fermeture peut être induite par des molécules lipophiles comme l’halothane. Un des rôles de ces jonctions est d’assurer le fonctionnement synchrone d’un ensemble de cellules d’un même type.
Communications par contact
Lorsqu’une cellule entre en contact avec une autre cellule (interaction cellule/cellule) ou avec la matrice extracellulaire (interaction cellule/matrice extracellulaire), elles peuvent adhérer l’une à l’autre et échanger des informations qui se traduisent par des manifestations détectables.
Les molécules qui interviennent dans ces interactions de type juxtacrine sont les « molécules d’adhésion ». On en distingue quatre grandes familles:
- les cadhérines, activées par le calcium, interviennent dans la morphogenèse et interagissent entre elles.
- les sélectines : protéines transmembranaires avec une partie intracytoplasmique et une partie extra-membranaire avec un NH2 terminal. La L-sélectine est constitutivement présente sur les leucocytes d’où elle peut être libérée dans la circulation par coupure protéolytique. La E-sélectine et la P-sélectine sont présentes à la surface des cellules endothéliales et plaquettaires activées.
- les intégrines: des glycoprotéines hétérodimériques sont exprimées à la surface de nombreuses cellules, lymphocytes, neutrophiles macrophages… Le LFA-1 (leucocyte function-associated molecule 1) et le VLA-4 (very late antigen 4) sont des intégrines.
- les immunoadhésines de la famille des immunoglobulines désignées sous le terme de CAM (cellular adhesion molecules) comprennent notamment :
- les ICAM (intercellular adhesion molecule) exprimées par les cellules endothéliales, les lymphocytes B et T, les monocytes, les fibroblastes, les cellules dendritiques…
- les PECAM (platelet-endothelial cell adhesion molecule) exprimées par les plaquettes, les cellules endothéliales, les lymphocytes.
- les VECAM (vascular endothelial cell molecule) exprimées par les cellules endothéliales.
